

文摘
孕產婦已知病毒感染風險的增加精神分裂症和自閉症的後代。使用這種觀察在動物模型中,我們發現懷孕小鼠呼吸道感染(包括BALB / c和C57BL / 6株)與人類流感病毒產生後代顯示高度不正常的成年後的行為反應。如精神分裂症和孤獨症,這些後代顯示赤字前脈衝抑製聲驚嚇反應(PPI)。還與控製老鼠相比,受感染的老鼠顯示驚人的反應急性抗精神病藥物管理局(氯氮平、氯丙嗪),擬精神病(氯胺酮)的藥物。此外,這些老鼠缺乏探索性行為開放田地和新奇物體的測試中,他們缺乏社會交往。至少其中一些行為變化可能是由於母體免疫反應本身。即孕產婦注入合成雙鏈RNA polyinosinic-polycytidylic酸導致後代PPI赤字沒有病毒。因此,產婦病毒感染對成年子女的行為有深遠的影響,可能通過一個胎兒母體的免疫反應的影響。
介紹
平均每年流感感染導致僅在美國10000 - 20000人死亡。此外,在過去的300年流感大流行發生每10 - 20年(陣風et al ., 2001)。流感感染的孕婦在懷孕會導致並發症如死胎、流產(該導彈和Glezen, 1994)。重要的是要注意,即使是不那麼嚴重產婦感染也會導致災難性的結果。許多流行病學研究發現精神分裂症的風險顯著增加女性的後代被暴露於流感在懷孕的第二個三個月。相似關聯了風疹、麻疹、水痘一帶狀皰疹和白喉(Mednick et al ., 1988;布朗et al ., 2000)。此外,產婦病毒感染被認為是“孤獨症的主要非遺傳性原因”(Ciaranello Ciaranello, 1995)。這些各種各樣的侮辱是什麼可能的共同點是孕產婦抗病毒反應(帕特森,2002)。
調查孕產婦病毒感染可能如何影響胎兒大腦發育,這將是有用的小鼠模型,使基因操作。幫助實現這個目標,我們報告一個初始的研究行為的呼吸道感染的老鼠的後代在mid-pregnancy mouse-adapted人類流感病毒。使用相同的孕產婦流感病毒感染的範例,我們之前發現薄的皮層和海馬,錐體細胞萎縮,減少水平的Reelin免疫反應性,改變神經元一氧化氮合酶的表達(nNOS)和synaptosome-associated蛋白質25 kDa (SNAP-25)和巨頭新生兒感染病毒的母親生出的小鼠的大腦中(銷et al ., 1995;法特米et al ., 1998 a,b,2000年,2002年)。
材料和方法
病毒感染。懷孕的9.5天,BALB / c小鼠(西蒙森實驗室、盡CA)麻醉腹腔內10毫克/公斤甲苯噻嗪和100毫克/公斤氯胺酮和注入經加州理工學院(Caltech) 6×103空斑形成單位的人類流感病毒A / NWS / 90年33 chiniμl PBS。Sham-infected母親相同的治療,但充滿了PBS。感染C57BL / 6小鼠(Simonson)同樣在猶他州州立大學(法特米et al ., 1999,2000年),母親和子女被運到加州理工學院行為分析,這是執行後4 - 8周的到來。後代是媽媽失散了3周後,和男性和女性在兩到四組分別關在籠子裏。
Polyinosinic-polycytidylic酸注入。Polyinosinic-polycytidylic酸(聚(我:C);σ,聖路易斯,密蘇裏州)在PBS稀釋,腹腔內注射在孕婦懷孕的雌性BALB / c 9.5天的劑量20日10日,5日,2.5,0毫克/公斤。
探索行為和焦慮。開放田地和新奇物體檢測的裝置包括一個50-cm-square開放透明的塑料盒子,17厘米。隻有25平方都在底部,和盒子被周圍熒光頂燈照明。在開放田地測試中,老鼠被放置單獨盒子的中心附近,他們的動作,隨後是在10分鍾內進行錄像。我們量化所花費的時間在9個中心廣場,廣場是進入的次數,hindpaws老鼠喂養的次數。使用相同的BALB / c小鼠(9個月),用於前脈衝抑製(PPI)下麵描述分析,一個觀察者忽視曆史的老鼠的這些行為。
新奇物體測試執行後立即開放田地測試沒有把老鼠從箱子裏。這樣做是為了條件之前鼠標打開”框模式將對象放置在箱子裏。對象是一個圓形的銀杯(直徑4厘米在封閉端,在開口端直徑6厘米,高4厘米),最後它被打開,從不管鼠標位於四個方塊,結束的時候開放田地測試,我們小心翼翼地把它放在一個正方形立即毗鄰一個平方襯砌牆的盒子。即對象總是在中心廣場但是不遠的牆上。聯係對象的延遲和聯係人的數量是估計的數量在最初的10分鍾。
自發的活動。運動活動是使用Photobeam活動監測係統(聖地亞哥儀器,聖地亞哥,CA)。一個半透明的籠子被photobeams之間。個別老鼠從家裏籠籠,和動態活動被定義為兩個連續中斷的四光束穿過籠40小時。
感覺運動協調。我們分析了老鼠rota-rod任務使用rota-rod跑步機(尤格Basile、Comerio、意大利)的光階段12小時光/暗周期。老鼠的位置時,計時器設置為零,rota-rod從4 rpm加速模式(增加4 rpm每25秒)。訓練後1 d的老鼠,我們進行了三個試驗對3 d,每天記錄最長的時間,每個鼠標rota-rod呆。這些時間平均為每個會話為每個鼠標,和方法計算的控製(n= 10)和暴露(n= 10)組。
社會行為。1到2 d開放田地試驗後,使用相同的開放田地盒子,我們研究社會互動對老鼠。我們放置兩隻老鼠被單獨安置)相同的性與實驗組相同盒子∼10厘米的中心和監控他們接觸另一個5分鍾的錄像測試在正常時期房間照明。積極聯係時打進一個鼠標的鼻子似乎< 2毫米遠離其他鼠標。接觸的時間包括任何類型的身體接觸。
聲驚嚇反應。在6 - 8周的年紀,代表青春期到成年早期,後期測試他們的後代在驚嚇聲驚嚇反應室(SR-LAB;聖地亞哥工具)使用前麵描述的標準方法(Swerdlow和蓋爾,1998年;科赫,1999)。5.1 -cm-diameter有機玻璃筒安裝在一個平台(20.4×12.7×0.4厘米)下麵附帶一個壓電加速度計單位與揚聲器位於sound-attenuated室(24厘米以上缸)和光。聲刺激的交付是由SR-LAB電腦控製的。噪音測量和校準無線電器材公司(沃斯堡,TX)音平計,和響應敏感性校準使用SR-LAB驚嚇校準係統。17分鍾測試會話由6個試驗類型:(1)脈衝(120分貝),40毫秒寬帶破裂;由20毫秒脈衝之前100 msec前脈衝(2)3 dB, (3) 5 dB,(4) 10分貝,對背景或(5)15分貝;(6)背景(65分貝)隻(不刺激)。測試會話後最初的5分鍾適應階段。我們的基線數據響應的強度和穩定性,習慣這些決定是一致的(Swerdlow和蓋爾,1998年)。
藥物治療,老鼠被放置在驚嚇框PPI檢測腹腔內注射後5分鍾。
結果
病毒感染
鼻內注入BALB / c和C57BL / 6小鼠6×103pfu人類流感病毒引起的疾病行為(嗜睡、困倦、折邊的皮毛和缺乏修飾)好幾天了,但失去了懷孕不是常見的,隻要老鼠沒有打擾。溫和(< 50%)肺癌合並發生在母親,與病毒效價的肺∼3 - 4×103空斑形成單位在感染的早期,效價下降之後到檢測不到的水平。發燒不發生在老鼠感染流感病毒(西德維爾et al ., 1986)。BALB / c的後代,出生體重的控製和實驗小鼠沒有明顯差異(分別為1.50±0.03 vs 1.34±0.09通用;p= 0.12;n= 9)。然而,垃圾的大小是不同的(8.25±1.03 vs 4.14±0.26 sham-infected vs感染病毒的母親,分別;p= 0.001;n= 7)。
探索行為
我們使用兩個探索性行為的化驗BALB / c小鼠,開放田地和新奇物體測試(文件,1985年;維斯et al ., 2000)。這兩個測試被認為是相關精神疾病的焦慮狀態。開放田地試驗,sham-infected與感染病毒的母親的後代很容易區分他們花的時間在中心廣場的盒子,他們進入那些方塊的次數。典型的行為的例子從兩組老鼠見圖1。定量的數據揭示了非常顯著差異在兩個探索性行為的措施(表1)。感染病毒的母親生出的小鼠花更少的時間在中心廣場(近8倍p< 0.0005;雙尾t測試),大部分時間都是花在盒子的角落(無花果。1)。這些老鼠也進入中心廣場近六倍少(p< 0.0016),和他們探討環境養育他們的後腿四倍少(p< 0.001)(表1)。這些發現在幾個較小的複製研究。

感染病毒的母親生出的小鼠的探索性行為非常不同於小鼠sham-infected母親所生。典型的鼠標的動作從每組從10分鍾的錄像帶會話跟蹤在一個開放的領域分為25平方線在底部的盒子。網站的飼養hindpaws說明了圈。雖然這些並不是最極端的例子從每組小鼠的行為,很明顯,鼠標生sham-infected母親(前)的反應非常不同的壓力,這種情況比鼠標一個感染病毒的母親所生底)。
雖然感染病毒的母親生出的小鼠10分鍾測試期間的大部分時間都在一個角落裏,他們沒有表現出凍結行為;他們經常移動。量化自發活動在籠子的環境中,我們使用一個photobeam-crossing裝置。當最初受感染的母親所生的老鼠被放置在這個新家裏籠子裏,他們清楚地顯示探索運動遠比控製老鼠(無花果。2)。這部小說雖然控製老鼠積極探索環境在第一個晚上,第一天實驗老鼠未能這樣做。然而,到了第二天晚上,回廊的數量在兩組小鼠運動是沒有區別的。因此,感染病毒的母親生出的小鼠的活動並不不同於控製一旦他們熟悉他們家籠裏的環境。這是一致的觀察實驗老鼠不是固定在開放田地測試;相反,他們經常但很少探索領域的中心(圖。1)。

自發活動由photobeam穿越化驗。個別老鼠從家裏籠和放在photobeam器40小時。連續梁的數量口岸(移動)每小時記錄和表達。控製老鼠(n= 8;4男性)探索這個新環境廣泛第一個24小時段,然後定居到第二天定期活動模式。感染病毒的母親生出的小鼠(n= 10;5男性)未能顯示早期,積極探索模式的控製,但是在第二個晚上開始,他們的基線活動水平沒有不同的控製措施。*p< 0.05;* *p< 0.01。
新奇物體測試執行後立即開放田地測試沒有把老鼠從箱子裏。兩項措施的對象探索被量化。感染病毒的母親生出的小鼠有幾乎兩倍大的延遲在第一次接觸對象(p< 0.05),他們開始近兩倍比老鼠更少的接觸對象(sham-infected母親生下的p< 0.003)(表1)。這些結果被複製在幾個小實驗。
感覺運動協調
測試老鼠一般感覺運動協調,我們使用的rota-rod加速模式。最初的訓練後,老鼠每天測試三次對3 d,我們平均每個鼠標的時間呆在酒吧在每個不同的會話(試驗1、2和3)在3 d;沒有明顯差異小鼠感染病毒的母親所生,出生sham-infected母親(圖。3)。

暴露和控製老鼠rota-rod測試顯示沒有差異。老鼠每天測試三次對3 d的rota-rod加速模式。結果表示為每個日常測試會話的方式,和我們持續時間最長的記錄,每個鼠標rota-rod呆。控製(沒有明顯區別n= 10)和暴露(n= 10)組織的任何三個交易日。
社會行為
使用相同的開放田地盒子,我們檢查了老鼠之間的社會互動,沒有見過彼此,但性與實驗組相同。感染病毒的母親生出的小鼠聯係彼此經常低於2.7倍的老鼠(sham-infected母親生下的p< 0.01)(表2)。也有四到五倍的不同延遲的第一次接觸,雖然這沒有達到統計學意義。大方差,在後者的數據集可能歸因於我們將另一個附近的老鼠開始測試,從而導致看似隨機接觸在某些情況下。
聲驚嚇反應
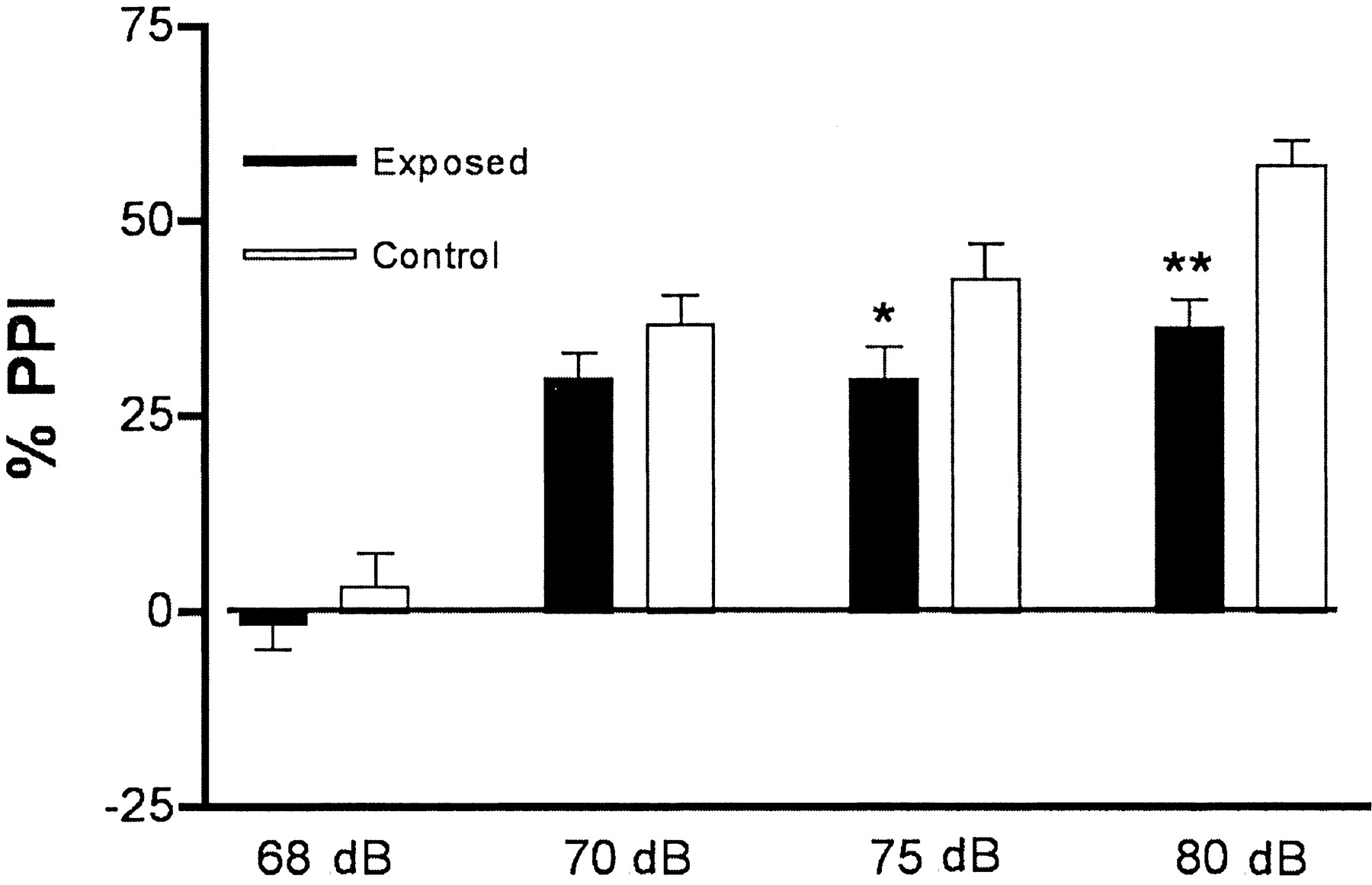
聲驚嚇反應已經被有效地用於衡量感覺運動控製的人類和齧齒動物。聲刺激了,驚嚇反應是以一個自動裝置。當一個prepulse-too造成驚嚇itself-precedes驚嚇刺激小,響應減少,這種現象稱為PPI。在一個實驗中使用相同的BALB / c小鼠進行探索性和社會行為,老鼠生sham-infected母親和老鼠感染病毒的母親還進行了PPI的誕生。各種前脈衝強度的PPI數據圖所示4。受感染的母親的後代顯示顯著的PPI赤字在75分貝的前脈衝(10 dB高於背景)和80分貝(15分貝高於背景)。相似的結果與C57BL / 6小鼠;生在一個實驗中,小鼠10 sham-infected母親和48小鼠感染病毒的母親所生進行測試,和後者顯示赤字在80分貝(數據沒有顯示)。

反應的BALB / c小鼠前脈衝抑製(PPI)分析。sham-infected母親出生的小鼠相比控製)(n= 14;8雌性),感染病毒的母親生出的小鼠(暴露)(n= 29;PPI 15雌性)顯示減少。在75分貝這些赤字是最重要的,這是10 dB高於背景(p< 0.05),在80分貝,這是15分貝高於背景(p< 0.01)。
使用精神藥物藥理測試已知的修改PPI進一步強調後代的兩組之間的差異。氯胺酮作為擬精神病代理,因為它的對立與穀氨酸NMDA受體加劇精神病症狀的精神分裂症患者在正常人和抒發幻覺(埃裏森,1995;Malhotra et al ., 1997)。像預期的那樣從先前的研究在大鼠(Swerdlow et al ., 19986毫克/公斤),注射氯胺酮導致控製減少PPI BALB / c小鼠(無花果。5)。相比之下,顯示的小鼠感染病毒的母親所生的實際增加PPI對氯胺酮的反應。驚人相似的結果被發現對C57BL / 6小鼠(無花果。5)。

氯胺酮在PPI的效果。擬精神病藥物氯胺酮是管理測試聲驚嚇反應前5分鍾。正如所料,氯胺酮減少PPI響應(80 dB前脈衝)BALB / c小鼠出生sham-infected母親(控製)(n= 14;8雌性),但它增加了PPI在老鼠身上感染病毒的母親所生(暴露)(n= 29;15雌性)。不同的實驗組和對照組之間的藥物引起的增加顯著(p< 0.02)。類似的結果觀察C57BL / 6小鼠(n= 10和48對實驗組和對照組,分別;p< 0.04)。
這些團體之間的區別的後代進一步突出顯示急性多巴胺受體阻滯劑。氯氮平、常用的非典型抗精神病藥物增加PPI齧齒動物(Swerdlow et al ., 1998),它是在我們控製BALB / c小鼠劑量依賴性的方式(數據沒有顯示)。然而,感染病毒的母親生出的小鼠顯示增加五倍大的PPI在回應2毫克/公斤氯氮平(無花果。6)。同樣的結果在C57BL / 6小鼠觀察(數據沒有顯示)。此外,非常相似的結果與兩株小鼠使用氯丙嗪2毫克/公斤,一個典型的抗精神病藥物(圖。6)。因此,抗精神病藥物糾正PPI赤字在實驗組,和這些老鼠顯示明顯對這些藥物過敏。

抗精神病藥物對PPI的效果。典型的抗精神病藥物氯丙嗪和非典型抗精神病藥物氯氮平敏銳地接種BALB / c小鼠,和PPI反應(80 dB前脈衝)化驗。如預期從此前的研究老鼠,老鼠sham-infected (控製)(n= 10;5雌性)顯示適度增加在PPI藥物注射後進行測試。相比之下,老鼠感染病毒的母親所生(暴露)(n= 21;12個雌性)顯示一個非常大的增加在PPI藥物管理局。藥物引起的差異增加了實驗組和對照組之間的(*具有重要意義p< 0.03)。注意,數據表示為PPI的比例增加。
這些藥理結果還表明,感染病毒的母親生出的小鼠不具有內在聽覺赤字,因為他們比控製老鼠的反應更強烈前脈衝在這種情況下。發現感染病毒的母親生出的小鼠顯示一個非常明確的PPI應對70年dB前脈衝,這是隻有5 dB以上(圖的背景噪音水平。4),表明小鼠正常聽力。
影響母體的免疫反應沒有病毒
這些結果所產生的一個基本問題是改變行為,我們觀察到受感染的母親的後代由於病毒感染的胎兒對母體的免疫反應或反應。此外,如果改變歸因於母體的免疫應答反應,會有抗病毒抗體引起病理學通過綁定一個抗原決定基之間共享病毒和胎兒的大腦?調查這個問題的一個方法是喚起一個antiviral-like母親不使用病毒免疫反應。這可以通過腹腔內注射的合成雙鏈RNA多聚(我:C)。保利(我:C)被喚起這種小鼠的免疫反應,包括upregulation主要組織相容性複合體的表達式,以及幹擾素等細胞因子(見參考內堂et al。(1999)和Verdijk et al。(1999)]。我們測試了幾個劑量(20、10、5,2.5,0毫克/公斤),如圖7,母親的後代注入劑量最高的聚(我:C)在PPI顯示赤字。這種赤字類似於病毒感染的母親的後代。因此,母體免疫反應,沒有病毒,就足以導致這種行為變化的後代。類似的結果在第二個、獨立實驗。

母體的免疫反應的影響沒有病毒。懷孕的BALB / c小鼠腹腔注射9.5天懷孕的三種劑量的聚(我:c)喚起一個antiviral-like免疫反應。PPI的後代測試4 dB前脈衝在6 - 8周的年齡層次。在最高劑量的聚(我:C),顯示的後代在PPI(*可觀的赤字p< 0.05;* *p< 0.01)(n為每個組= 10;5每個女性)。
討論
我們的數據對小鼠流感病毒感染母親所生顯示高度的顯著變化行為開放田地,新鮮事物和社會交互測試。這些行為的變化可能反映了小說焦躁不安或緊張的情況下,這是一個自閉症的突出特點。在處理壓力在精神分裂症中也發現是困難。此外,這些老鼠顯示赤字PPI和非常獨特的PPI對急性擬精神病和抗精神病藥物的反應。赤字在PPI中觀察到一些精神疾病,包括精神分裂症(蓋爾et al ., 1999和自閉症McAlonan et al ., 2002)。明顯異常反應擬精神病和抗精神病藥物,我們發現這些老鼠建議改變多巴胺能和glutamatergic係統的可能性。精神分裂症的當前理論假定這兩個係統的不平衡;這種不平衡在這些老鼠可能會造成他們的藥物反應。最近的工作也已經確定了自閉症的穀氨酸神經遞質係統的變化(珀塞爾et al ., 2001)。因此,感染病毒的母親生出的小鼠的行為符合預期的齧齒動物版本的精神分裂症和自閉症。所有的測試是特定用於這兩種障礙,然而。有幾個其他病毒感染模型,主要利用產後老鼠,這也顯示明顯的異常行為(審查,請參閱何寧,利普金,2001;皮爾斯,2001;帕特森,2002)。
罷工的感染病毒的母親生出的小鼠的行為變化表明,大腦發育改變了這個胎兒環境的擾動。使用相同的孕產婦流感病毒感染的範例,我們之前發現幾個大腦組織學的變化(銷et al ., 1995;法特米et al ., 1999)。這將是有趣的,以確定哪些這些(或其他)分子和形態改變與最好的PPI赤字在實驗老鼠中找到。這樣的研究可以揭示大腦回路以及分子參與正常的PPI和聲驚嚇反應,通過類比,自閉症和精神分裂症的變化主題展覽PPI異常。
更大的問題是:病毒感染的母親如何影響胎兒的大腦發育呢?我們懷疑不太可能實際上流感病毒感染胎兒的大腦,因為限製的非致命性的病毒株呼吸道最常見,其中的關鍵細胞因子響應的分辨率感染(威廉姆斯和麥肯齊,1977年;歐文et al ., 2000;湯佩使et al ., 2000;範Reeth 2000)。雖然我們使用的應變可以親神經的(Stuart-Harris et al ., 1985),我們無法檢測到病毒在受感染的母親所生新生兒的老鼠的大腦用空斑實驗和培養的腦組織(r·w·西德維爾,未發表的觀察)。測試抗病毒抗體染色的大腦是負的,和沒有出現腦炎部分從這些新生兒大腦(未發表的觀察,s·h·法特米)。此外,初步rt - pcr檢測未能檢測到病毒RNA從胎兒的大腦從受感染的母親(施n, l ., p h·帕特森,未發表的觀察)。
或許更重要的是,我們發現在母親的後代PPI赤字注射合成雙鏈RNA多聚(我:C),喚起一個antiviral-like小鼠的免疫反應。這表明母體免疫反應就足以導致成年子女的行為改變,至少在PPI。胎兒大腦發育的潛在效應物改變在這個模型中就包括糖皮質激素和細胞因子。這些可能性是服從實驗操作。這些數據可以用來測試治療操作改善或預防流感病毒感染的有害影響胎兒的大腦發育。的證據(引言中提到的)孕產婦病毒感染會增加精神分裂症和自閉症的風險,有趣的是,各種免疫異常在精神分裂症已報告(賴特和穆雷,1993年;穆勒et al ., 1999;那霸et al ., 2000;保盈et al ., 2001;Rothermundt et al ., 2001和自閉症漢堡和沃倫,1998年;Jyonouchi et al ., 2001)。
我們選擇了9.5天感染在這項研究中,因為它是大約mid-pregnancy鼠標,和人類流行病學研究強調懷孕中期作為流感的關鍵時刻作為一個風險因素。這個階段在鼠標恰逢神經脊遷移到臉和中腦和時間Cajal-Retzius細胞產生Reelin。Reelin是重要的成神經細胞遷移,其表達異常在精神分裂症的大腦(Impagnatiello et al ., 1998;法特米et al ., 1999),這是符合這個障礙(的發展理論Marenco溫伯格,2000;那霸et al ., 2000;托裏和Yolken一起,2000)。鑒於幾部分老鼠大腦的發展晚於人類,老鼠母親懷孕的後期感染的研究可以揭示同樣有趣的結果。
除了這個工作的相關精神疾病,調查潛在的治療方法是很重要的,因為母親流感感染引起的嚴重問題。這種感染可能對胎兒造成嚴重的後果,包括流產、早產、死胎,和早期新生兒死亡率,還有一個母親死亡率的風險增加(該導彈和Glezen, 1994)。
腳注
這項工作是由薑和泰德·詹金斯的禮物和梅特勒自閉症格蘭特P.H.P. S.H.F.菲利斯和佩裏Schwartz研究國家聯盟精神分裂症和抑鬱症的調查員。R.W.S.支持契約式n01病毒學分支- ai - 65291,國家過敏症和傳染病研究所,國立衛生研究院。我們感謝d·麥克道爾和f .白嘴鴉行政幫助,j·貝爾關於老鼠的幫助,和l . Tecott行為測試的建議。
信件應該寫給·h·帕特森的生物學216 - 76年,加州理工學院的帕薩迪納,CA 91125。電子郵件:php在}{caltech.edu。








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}